
A bird’s brain is similar to that of mammal’s but there are difference in the acuity of the senses, especially sight. In addition, the cerebrum, the “thinking ” part of the brain is not as well developed because birds are more instinctive than learners. But their cerebellum, the center of mechanical coordination, is well developed for flight.
SIGHT
Birds are highly visual animals; they have to be to be able to fly. The importance of birds’ eyes is implied by their size – they are the largest relative to the body of all animals. Some hawks and owls have eyes as large as human eyes. The eyes of the ostrich, at 50 mm in diameter are the largest of any land vertebrate. In some owls, the eyes comprise up to 1/3 of the total weight of the head. In the Starling, the eyes comprise 15% of the head weight; in humans it is only about 1%. In most aspects, the avian eye structure resembles that of mammals.
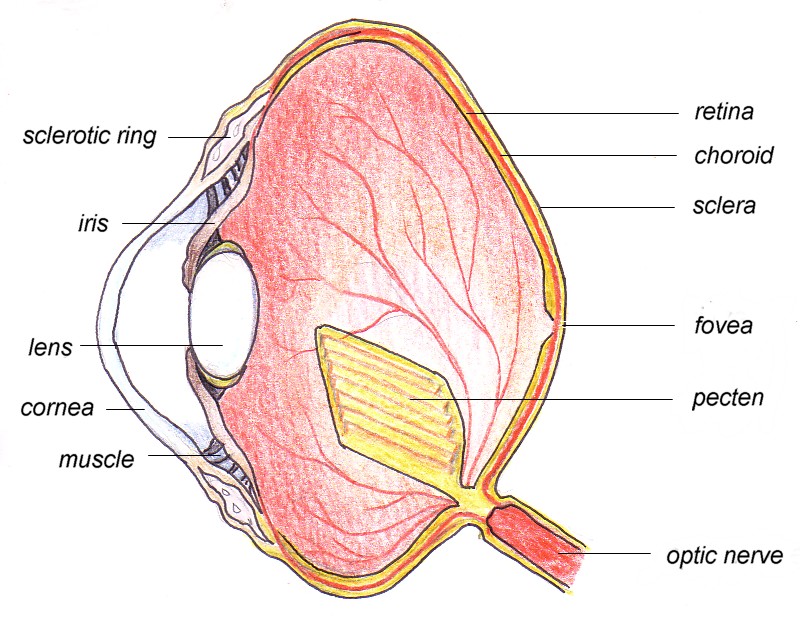
The eyes of a bird are able to adjust to the light about 2x as well as that of a 20 year old human. The lens is very flexible in most birds and can change shape readily – birds need to focus near and far and change rapidly. The entire eye also varies in shape form flat to round to tubular (hawks and owl); the shape is maintained by a ring of overlapping bony plates.
The retina is the sensitive layer of the eye that absorbs the incoming light, senses it, integrates the information in it, and sends this information to the brain. The entire retina is thicker than that of mammals and the rods and cones are more abundant. Cones are for color and rods are for black and white and dim light. The distribution and density and proportion of rods to cones varies with the species of bird. Diurnal birds have retinas dominated by cones while nocturnal birds have mostly rods. Many hawks and owls have more sensory cells in the upper half of the eye that receives more light when the birds look down.Color vision varies among birds; some, but not all, see color. Hummingbirds, like insects, can see ultraviolet light.
Any part of the retina that has a denser concentration of receptor cells perceives sharper images. One such area of densely packed cells is called a fovea. Not only are the cells denser in that area but the pit shape serves to refract the light so that a larger image is formed. Humans have a fovea; when you focus directly on an object the image falls on the fovea- that’s why peripheral vision is weak. Overlapping of the visual fields of the two eyes produces binocular vision and depth perception. Most birds have a one-fovea eye that functions similarly to ours, but many birds are bifoveal. One fovea birds have the fovea located near the place where the optic nerve enters the eye- a central fovea. In the birds with two foveas, especially those that need to be good judges of speed and distance, the second fovea is located in the temporal region of the eye. This is typical of birds of open country- hawks, eagles, terns, parrots, swallows, doves. This additional temporal fovea broadens the width of sharp focus and helps to judge speed and distance; e.g.’ hawks, terns, parrots, swifts, and hummingbirds. A few birds even have a third fovea – a few terns and swallows. A few other species of birds have only one poorly developed fovea – owls and sandpipers and thus bob their head to gain perspective.
There has been a lot of debate regarding the acuteness of avian vision. It appears that it is better than human vision but perhaps not significantly so. A vulture sees about as sharply as humans but a chicken appears to see about 1/25th as well as humans and the hawks and songbirds see about 21/2 times as sharply. But with a double fovea, birds can keep track of a moving object easier than can a human. Pigeons, e.g., can detect movement as slow as 15 degrees per hour.
The density of receptor cells in the retina also allows birds to see in very dim light. Barn Owls can see an object at 2 meters with an illumination of .00000073 foot candles. This is equivalent to a person seeing an object by the light of a match a mile away.
The nictitating membrane (third eyelid) is used for blinking; eyelids are used only for sleeping. It is covered with brushlike processes inside so that the cornea is swept with tears every time the membrane moves over the eye. In diving birds this membrane serves as a contact lens. In owls, unlike other birds, it is opaque. In the Black-billed Magpie, this third eyelid has an orange spot that is displayed during courtship or aggressive behaviors. It is also found in some amphibians, reptiles, and some mammals. In some diving birds such as loons and grebes and ducks and auks the nictitating membrane has a clear center which acts as a contact lens under water.
HEARING
Birds’ ears are divided into three parts -external , middle, and inner.
In terrestrial vertebrates the organ of hearing probably first evolved as an organ of equilibrium and later as one for hearing. In reptiles and birds a single bone transmits sound vibrations from the eardrum to the cochlea while in mammals, we have the malleus, incus, and stapes.
In most birds, specialized feathers surround the ear opening (except ostriches, vultures, and some galliformes) to minimize air turbulence. The shape of the outer ear varies; the shape of the ear openings also differ. The most extensive variations of the ear are found among the owls- many species have evolved very large ear flaps which look almost like human ears. In several owls the ears are bilaterally asymmetrical (unique among vertebrates) to helplocalize sound.
Owls can detect prey in total darkness with an error of only 10 horizontal and vertical. Owls can also change the position of their auricular feathers and to some extent the shape of their ear. The heart-shaped face of the owl helps collect sound and direct it to the ears.
The cochlea transmits vibrations to the auditory nerve. There seems to be a correlation between the length and complexity of the cochlea and complexity of the song of that species.

The frequency range of sound waves a single bird species can receive is narrower than that of mammals. Birds are less sensitive to the high and low ends of their range than mammals, but in the middle frequencies it is similar. However, birds are about 10x as sensitive to rapid fluctuations in pitch and intensity than humans. Birds can distinguish between frequencies that differ by 1% or less and they can distinguish between sounds separated in time by only .6 to 2.5 milliseconds.
A few birds use echolocation, like bats. The Oilbird and the Cave Swiftlet of southeast Asia are examples. They use echolocation to navigate but not to catch prey. Recent evidence indicates, however, that penguins can locate prey via echolocation.
Both Darwin and Audubon showed that vultures did not find carcasses if they were covered. Audubon painted a picture of a dead sheep and vultures tried to eat it. There is also an anecdote about Black Vultures attacking and eating a spraying skunk. And migrating vultures have been disoriented by releasing smells into the air.
Birds have been tested for their ability to smell by training them to discriminate between odorless and odorous air; e.g. pigeons were trained to peck at a disc when they smelled an odor. Vultures can smell carrion because carrion emits the gas ethyl mercaptan; some enterprising engineers put ethyl mercaptan in a gas line to detect leaks and found them when they saw vultures circling over a part of the gas line.
Albatrosses, skuas, and petrels in the Antarctic seem to be able to detect meat, fat, or blood spread on the sea’s surface. Leach’s Petrel do some navigating by olfactory clues – they can find their nests at night by flying from downwind into the wind coming from the colony and find their own nesting material even if moved to a different burrow.
Evidence indicates that African Honeyguides can locate beehives by the odor of the wax. Homing pigeons may also use olfactory clues for navigation; birds with experimentally plugged nostrils took longer to find their way home. The flightless Kiwi is nocturnal and feeds on food hidden in the soil. Its vision is poor and it is the only bird with nostrils at the end of the beak and sniffs while foraging.
In general, olfactory senses of birds are poor, but there are exceptions and we might just think they are poor because we don’t have evidence.
TASTE
Taste buds, concentrated on the posterior part of the tongue and pharynx floor, are similar to mammalian ones. The total number of taste buds is less, though.
Sweet– Many birds show little interest in sugar/sweet taste except for parrots, hummingbirds, and other nectar and fruit feeders. Hummingbirds can distinguish different kinds of sugars and their concentrations. Seed eaters show no preference for sweet or sour.
Salt – For most birds there is a variety of salt discrimination levels. For sea birds tolerance for salt water is high; they can excrete excess salt through their nasal glands although they will drink fresh water in preference to salt water if given the choice.
Sour– Birds have a wide range of tolerance for sour.
Bitter– Again, a variety of responses. The classic study was done by Brower in 1969. Many species of milkweeds contain cardiac glycosides. If the milkweed is eaten by an animal the heart rate drops but the beat is stronger; a large enough dose is fatal. But 1/2 the fatal dose causes vomiting so an animal will throw up before it absorbs a lethal dose. Many animals learn, then, not to eat milkweeds. Some insects, however, can eat the plants with no apparent side effects. One is the caterpillar of the Monarch Butterfly. Brower raised Monarchs in captivity; some on milkweed and some on cabbage, which has no cardiac glycosides. The cabbage-raised butterflies were fed to Blue Jays and they ate them with no ill effects. Then the birds were offered the monarchs raised on milkweed; 12 minutes later the Blue Jays became violently ill, vomited, but recovered in 1/2 hour. Then the Blue Jays would reject all Monarchs introduced into the cages, no matter how they were raised. If starved, the Blue Jays would capture and pick at the butterflies to taste them first.
The point of this is that birds can taste bitter and will learn to associate bitter with plants or animals and avoid them. Both the predator and prey benefit as the prey doesn’t get eaten and the predator doesn’t get sick and waste time eating them.
TOUCH
A bird’s skin possess sensory nerve endings, as in mammals, that detect heat, cold, pressure, and pain. Many are also associated with filoplumes. The tip of the bill of ducks and geese, the tongues of woodpeckers, and the bill tips of many altricial birds have concentrated nerve endings. In many shorebirds there are pits with sensory cells along the bill that are very sensitive to pressure – for finding prey. Woodstorks, for instance, can fish in turbid water and close their beak on live fish in .019 seconds after initial contact (takes us .040 seconds to blink our eyes). Similar sensory cells are also found at the base of flight feathers and it is likely that they play an important sensory role in flight.
CENTRAL NERVOUS SYSTEM
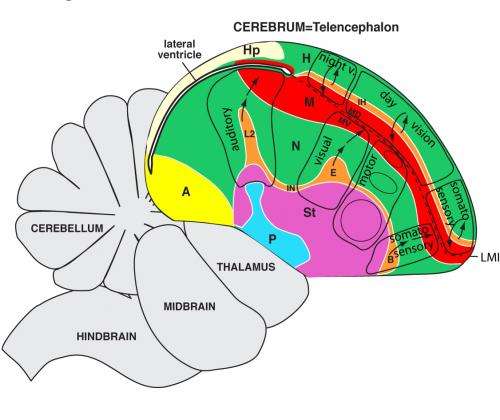
The central nervous system (brain and spinal cord) is arranged and works similarly to that of mammals. Its job is to integrate sensory impulses from the environment, to stored learned information, and to coordinate voluntary and involuntary functions and movements.
In the brain, the optic lobes are large and the olfactory lobes small, correlating with their senses.
The cerebral hemispheres in birds are large and well-developed, as in mammals, but the center of complex behavior in the cerebrum is different in the two groups. The brain of a mammal is dominated by the top layer of the cerebral hemispheres (cerebral cortex) which have a high capacity for learning. The bird brain is dominated by the middle of the cerebral hemisphere which lacks learning capacity. So mammals, in general, learn behavior and bird behavior tends to be instinctive and stereotyped. Birds’ brains are also particularly sensitive to control by hormones; implanting a pellet of testosterone in a dove elicits courtship, copulation, and aggressive behavior. Experimentally, large sections of the cerebral cortex have been removed with little effect on the birds’ behavior.
The cerebellum, the center for motor control, is well-developed in birds as is logical with their ability to fly and the related need for agility.
hello, I am a 5th grader at stem academy in fond du lac Wisconsin, I am curantly doing a project on hawks, because I have a sharp shinned hawk, living very near me. I would like to know more about the CENTRAL NERVOUS SYSTEM so that I can meet my standard. If you could reply to this I would very much love that. I do not have a web site, so I put in my email!!!!
You can read about the central nervous system in an anatomy or ornithology book. It is too much for me to try and explain it here.
i heard that humans can only use half of their true muscle strength because our brains limit how much strength we can use. Do the brains of birds also limit their strength in the same way?
I have never heard of either humans or birds brains controlling muscle strength but it makes some sense. But bird brains are much more about muscle control and coordination than humans are. Could make a difference.
Hey,
I was exactly searching for the working of the nervous system of animals, and your article is worth reading.
thank you.
Pingback: Can Birds See at Night? Everything You Need to Know - Bird Watching Pro
Can birds lose their sight too just like humans?
Yes, disease or accident. A fatal flaw.
Things about birds we always wondered!
Excellent web site!